


Isolation and genetic analysis of the chikungunya virus from Aedes aegypti and Aedes albopictus mosquitoes captured in Central America
- Authors: Ignatyev G.M.1, Oksanich A.S.1, Kazakova E.V.2, Samartseva T.G.1, Otrashevskaya Е.V.2, Uyba S.V.3, Trukhin V.P.2
-
Affiliations:
- I. Mechnikov Research Institute for Vaccines and Sera
- St. Petersburg Research Institute for Vaccines and Sera
- Mechnikov Latin American Institute of Biotechnology
- Issue: Vol 100, No 5 (2023)
- Pages: 310-318
- Section: ORIGINAL RESEARCHES
- URL: https://microbiol.crie.ru/jour/article/view/18477
- DOI: https://doi.org/10.36233/0372-9311-354
- EDN: https://elibrary.ru/uwguml
- ID: 18477

Cite item

Abstract
Introduction. The habitat of mosquitoes belonging to the genera Aedes spp., Culex spp., Culiseta spp. is in South and Central America, including Nicaragua. Monitoring of the spread of mosquito vectors and assessment of the infection with arboviruses can provide information on possible occurrence of new diseases or an increase in the reported cases, changes in the infectivity of viruses for humans due to changes in pathogen transmitters.
The purpose of this study was isolation and identification of arboviruses belonging to the Flavivirus and Alphavirus genera from A. albopictus, A. aegypti, Culiseta spp., Culex spp. mosquitoes captured in forests of Nicaragua.
Materials and methods. A. albopictus, A. aegypti, Culiseta spp., Culex spp. mosquitoes were captured during the dry season in 2021 in forested areas of Nicaragua in four different locations. Mosquitoes were sorted into pools, each containing 5-8 mosquitoes (236 pools in total). Using the reverse transcription polymerase chain reaction, the pools were tested for the presence of chikungunya (CHIKV), dengue, Zika, and yellow fever viruses. Positive pools were inoculated into the C6/36 cell culture to obtain isolates and for their further sequencing.
Results. The dengue virus was detected only in Aedes spp. mosquitoes: in 7 pools — A. aegypti, in 1 — A. albopictus. CHIKV was also detected only in Aedes spp. mosquitoes: in 3 pools — A. aegypti, in 1 — A. albopictus. The sequencing of nucleotide sequences of 6К, Е1, Е2, and NS1 genes of CHIKV isolated from A. albopictus mosquitoes showed that compared to the similar gene sequences from CHIKV isolates recovered from A. aegypti mosquitoes, the 6K gene region contained 4 nucleotide and 4 amino acid substitutions, while the E1 region contained 16 nucleotide substitutions, 10 of them led to amino acid substitutions; the E2 region contained 14 nucleotide and 11 amino acid substitutions; the NS1 region contained 33 nucleotide and 19 amino acid substitutions.
Full Text
Introduction
Mosquitoes of the genera Aedes spp., Culex spp., Culiseta spp. are vectors for numerous pathogens causing viral diseases such as dengue fever, Zika fever, yellow fever, chikungunya fever, Venezuelian equine encephalomyelitis, Sindbis fever and belong to families Flaviviridae (genus Flavivirus) and Togaviridae (genus Alphavirus) [1–6]. Habitats of mosquitoes belonging to the genera Aedes spp., Culex spp., Culiseta spp. are located in South and Central America, including Nicaragua [2, 3, 7–10]. The monitoring of the spread of mosquito vectors and the assessment of their infectivity can provide information on possible occurrence of new diseases or an increase in the reported cases, while the change of a pathogen’s vector can cause changes in the infectivity of viruses for humans [5]. Isolation of viruses directly from vectors captured in their natural habitats and exploration of isolated strains are essential for development of diagnostic tools, prevention and treatment of diseases caused by alphaviruses and flaviviruses.
The purpose of this study was isolation and identification of arboviruses belonging to the genera Flavivirus and Alphavirus from A. albopictus, A. aegypti, Culiseta spp., Culex spp. mosquitoes captured in forests of Nicaragua.
Materials and methods
Mosquitoes
Mosquitoes were captured during the dry season in 2021 in a forested area of Nicaragua in four different locations having the following coordinates:
- location 1 — 12.325527N 85.974662W;
- location 2 — 12.323326N 85.974275W;
- location 3 — 11.908210N 85.932490W;
- location 4 — 11.903555N 85.938758W.
Representatives of the genera Aedes spp. (A. albopictus, A. aegypti), Culiseta spp., and Culex spp. were among the captured mosquitoes. After their species had been identified, mosquitoes were sorted into pools of 5–8 insects of the same species captured in the same location. A total of 236 pools were formed. Each pool was homogenized in 300 μl of the Leibovitz L-15 medium (Gibco, Thermo Fisher Scientific), pH 7.4, using ceramic beads and the SpeedMill Plus homogenizer (Analytik Jena) to obtain the suspension. 140 μl of homogenate from each pool was used for RNA extraction, which was performed using a QIAamp Viral RNA reagent kit (Qiagen). The reverse transcription polymerase chain reaction (RT-PCR) was performed using protocols for each pathogen. The pools tested positive for any of the viruses under study were used for subsequent isolation of the virus on cell cultures.
Virus isolation
The remaining mosquito suspension (200 μl) was filtered through the 0.45 µm PES filter. The obtained filtrate was used to infect the monolayer of C6/36 cells grown in 24-well plates. The inoculation was performed in 100 μl using the stock and dilution 1:10. Seven days after the infection, 100 μl of the supernatant were used for the next passage. A total of 5 successive passages were conducted. After the infection, the cells were examined daily for the presence of any signs of a cytopathic effect.
Molecular and genetic study
The RT-PCR-based molecular and genetic study was performed using the following procedure. The 140 μl mosquito suspension was used to extract RNA using a QIAamp Viral RNA reagent kit (Qiagen) in accordance with the instructions of the manufacturer. Then using the reverse primer pNS1CHVrev2-3 for the chikungunya virus (CHIKV; the NS1 gene region); the mixture of reverse primers for the dengue virus panDVrev1 and panDVrev2 (the 3’-UTR region), which were common for all 4 types of the virus (Table 1); the reverse primer for the Zika virus pZVrev [9]; the reverse primer for yellow fever virus pYFVrev (Table 2) [11] and a reagent kit for RT (Syntol), the cDNA was generated from the viral RNA template using an RT reaction. During the first stage, 2 μl of reverse primers (10 pmol/μl) were mixed with 6 μl of the extracted RNA; then, the mixture was heated at 95оC for 5 min. Then, the tubes were cooled down at the room temperature for 2 min and 22 μl of the mixture were added for RT (9 μl of deionized water, 12 μl of the 2.5-fold buffer for RT (Syntol), 1 μl of MMLV-reverse transcriptase (Syntol)) and were incubated at 42оC for 30 min. The mixture was heated at 95оC for 5 min to inactivate the reverse transcriptase.
Table 1. Nucleotide sequences of primers and probes for detection of CHIKV and dengue virus RNA in collected samples using real-time RT-PCR
Oligonucleotide | Nucleotide sequence, 5’–3’ | PCR product size |
CHIKV (the NS1 gene region) | ||
pNS1CHVfor | GTGTGCTGTTCTCAGTAGGGTCAACG | 218 bp |
pNS1CHVrev | GTCTGCGTGGTGGGTTACCGC | |
zNS1CHVfor | FAM-GGCTACGTCGTTAAGAGAATAACGATGAGCCC-BHQ1 | |
Dengue virus (the 3’-UTR region) | ||
panDVfor | GACTAGYGGTTAGAGGAGACCC | 190 bp |
panDVrev1 | CGTTCTGTGCCTGGAATGATG | |
panDVrev2 | CGCTCTGTGCCTGGATTGATG | |
zDVfor | FAM-GCATATTGACGCTGGGARAGACCAGAG-BHQ1 | |
Note. Y — either C or T; R — either A or G.
Table 2. Nucleotide sequences of primers for amplification of Zika and yellow fever virus RNA using RT-PCR and electrophoresis detection
Oligonucleotide | Nucleotide sequence, 5’–3’ | PCR product size |
Zika (the NS5 gene region) | ||
pZVfor | CCGCGCCATCTGGTATATGT | 450 bp |
pZVrev | CTCCACTGACTGCCATTCGT | |
Yellow fever (the E gene region) | ||
pYFVfor | TACCCTGGAGCAAGACAAGT | 465 bp |
pYFVrev | GCTTTTCCATACCCAATGAA | |
For the Zika virus and yellow fever virus, agarose gel electrophoresis detection of PCR amplicons was performed using non-original primers from other researchers [9, 11], which are listed in Table 2. The amplification was performed in accordance with the following program: 95оC — 1 min 30 sec; 30 cycles: 95оC — 20 sec, 55оC — 15 sec, 72оC — 30 sec; 72оC — 10 min.
The real-time PCR was performed for CHIKV and dengue viruses using original primers and probes. The DTprime instrument (DNA-Technology) was used for amplification following the program: 95оC — 1 min 30 sec; 40 cycles: 95оC – 15 sec, 55оC – 40 sec.
The in-house designed oligonucleotides and the procedure described previously were used for generating CHIKV 6К, E1, Е2, and NS1 gene fragments and their sequencing [12].
Prior to sequencing, all the PCR products were gel-purified using a Cleanup Standard reagent kit (#BC022, Evrogen) and cloned the pGEM-T Easy vector (Promega) in accordance with the manufacturer’s instructions. Then, the clones were selected, plasmids were extracted, and the Sanger sequencing was performed using standard primers for sequencing T7 and SP6. The obtained nucleotide sequences were aligned using the MEGA11 software1.
Phylogenetic analysis
The phylogenetic analysis was conducted using the molecular dating method, the BEAST v. 1.10.4 software package and the associated software suite BEAUti v. 1.10.42. When constructing trees, we used the HKY3 nucleotide substitution algorithm with a strict molecular clock. The analysis included 10 million trees, every 1,000th tree was selected. The MEGA11 software was used for tree construction. The multiple alignment was performed using genetic sequences of genome fragments (E2, 6K, E1) of CHIKVs isolated from mosquitoes (30 sequences) in different regions of the world. The position in the genome fragment, which was used for the phylogenetic analysis, was 8574–11 303 bp (the positions are shown for the prototype strain isolated in 1959, GenBank KX262990). The phylogenetic tree was extended with 8 nucleotide sequences of CHIKV isolated from humans, 3 of which were obtained from patients in Nicaragua in 2014 and 2015, and 4 of them were reference sequences of the virus.
The infectivity of mosquitoes was measured following the previously described procedure [10, 13].
Results and discussion
The mosquitoes captured in four locations represented three genera Aedes spp., Culex spp., Culiseta spp. As can be seen from Table 3, the best represented genus was Aedes spp. Accounting for 954 mosquitoes: 604 (67%) mosquitoes belonged to A. aegypti, 314 (33%) mosquitoes belonged to A. albopictus. Out of 105 pools of A. aegypti mosquitoes, 7 pools were tested positive for the dengue virus RNA, while 3 pools were tested positive for CHIKV RNA. Zika virus and yellow fever virus RNAs were not detected. No simultaneous detection of CHIKV RNA and dengue virus RNA was recorded. Out of 54 pools of A. albopictus mosquitoes, the dengue virus RNA was detected in 1 pool and the CHIKV RNA was also detected in 1 pool. Zika virus and yellow fever virus RNAs were also not detected. The simultaneous detection of CHIKV and dengue virus RNA was not reported. No RNA of the tested viruses was detected in 46 pools of mosquitoes belonging to the genus Culex spp. and in 31 pools of mosquitoes representing the genus Culiseta spp.
Table 3. Genus and number of captured mosquitoes
Mosquito species | Number of mosquitoes | Number of pools | PCR results (positive/negative) | |||
dengue virus | CHIKV | yellow fever virus | Zika virus | |||
Aedes spp. | 954 | 159 | 8/151 | 4/155 | 0/159 | 0/159 |
A. aegypti | 640 | 105 | 7/98 | 3/102 | 0/105 | 0/105 |
A. albopictus | 314 | 54 | 1/53 | 1/53 | 0/54 | 0/54 |
Culex spp. | 278 | 46 | 0/46 | 0/46 | 0/46 | 0/46 |
Culiseta spp. | 188 | 31 | 0/31 | 0/31 | 0/31 | 0/31 |
- albopictus mosquitoes from the CHIKV positive pool were captured in location 1 and A. aegypti mosquitoes from the positive pools were captured in location 4. The dengue virus RNA was detected in pools of A. aegypti mosquitoes captured in location 2, and Aedes albopictus mosquitoes captured in location 2.
The minimum infection rate for A. aegypti mosquitoes for the dengue virus was 10.0 and for CHIKV – 4.6. The minimum infection rate for A. albopictus mosquitoes for the dengue virus and CHIKV was 3.0. The obtained results lead to the assumption that mosquitoes of the genus Aedes spp. were infected with the dengue virus and CHIKV.
The minimum infection rate for mosquitoes, undoubtedly, is an important variable; however, it depends on the number of captured mosquitoes (the size of the studied subset) and the number of mosquitoes in the pool, thus affecting the sensitivity of the employed method. In some studies of the minimum infection rate among mosquitoes infected with alphaviruses and flaviviruses, the rate ranged from 0 to 12, thus implying a potential risk of infection with this virus transmitted by the particular mosquitoes [10, 13]. The possible coinfection with dengue and chikungunya viruses was described earlier; the examination of serological markers of flaviviruses and alphaviruses demonstrated the presence of antibodies against the dengue virus and CHIKV [9, 14]. The RT-PCR positive mosquito pools – 8 pools with the dengue virus and 4 pools with CHIKV were used to isolate viruses.
C6/36 cells were used to isolate viruses from the mosquito pools. Each pool was put through 5 successive passages. In the cells infected with samples containing CHIKV (based on the RT-PCR results), the disturbance of the cell monolayer – the development of a cytopathic effect – was observed in the 2nd – 3rd passages. In the 5th passage, the 100% cytopathic effect was observed after 72 hours. In the samples containing dengue viruses (based on the PCR results), the cytopathic effect was not as pronounced. In each passage, the authenticity of the isolates and the absence of cross-contamination were verified using RT-PCR. After the isolation had been completed, the isolates were sequenced. The resulting isolates contained CHIKV and dengue viruses types 1 and 2. In isolates containing dengue viruses, only the type of the virus was identified due to the short length of PCR product fragments.
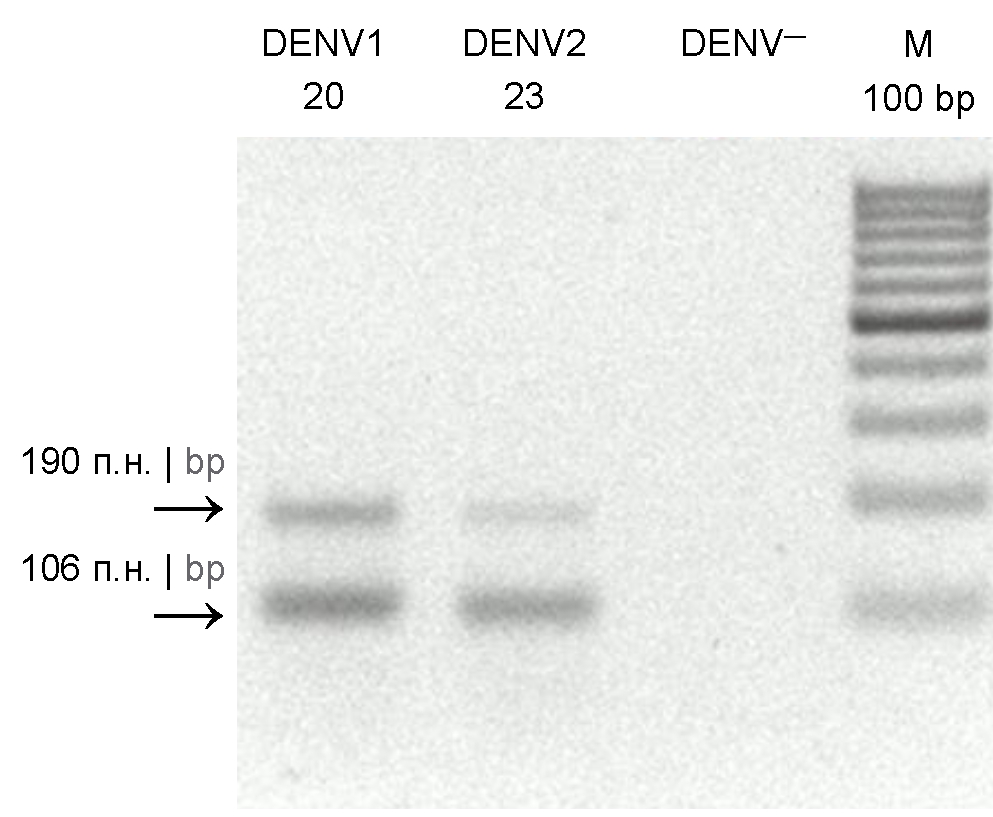
As can be seen from the electropherogram of amplicons (Fig. 1) obtained using universal primers for all the 4 types of the dengue virus, the amplification produces 2 amplicons 190 and 106 bp long. The sequencing of fragments showed that both of them were specific [15, 16].

Fig. 1. Agarose gel electrophoresis (2%) of PCR products. DENV1 20 and DENV2 23 — amplicons of dengue viruses types 1 and 2 from 20 and 23 pools, respectively; DENV— — negative PCR controls for the dengue virus; M 100 bp – DNA molecular weight marker.
The sequencing of nucleotide sequences of CHIKV 6К, Е1, Е2, and NS1 genes from A. albopictus mosquitoes (1 pool, location 1) showed that it was the same strain, which we had isolated in the same location in 2018 [12]. Compared to the similar gene sequences from the CHIKV isolates obtained from A. aegypti mosquitoes (3 pools, location 4), the 6K gene region had 4 nucleotide and 4 amino acid substitutions; the E1 region had 16 nucleotide substitutions, 10 of which resulted in amino acid substitutions; the E2 region had 14 nucleotide and 11 amino acid substitutions and the NS1 region had 33 nucleotide and 19 amino acid substitutions (Table 4). Three isolates from A. aegypti mosquitoes did not have any amino acid substitutions relative to each other. The isolated strain was deposited in the genetic sequence database of GenBank (NCBI) with accession number OQ320495.
Table 4. Nucleotide and amino acid substitutions in 6K, E1, E2, and NS1 gene regions in CHIKV isolates recovered from A. albopictus and A. aegypti mosquitoes captured in different locations in Nicaragua
Substitutions A. albopictus > A. aegypti | |||||||
6K protein gene | E1 protein gene | E2 protein gene | NS1 protein gene | ||||
N | А | N | А | N | А | N | А |
A37G A54T C107T T176C | E13K F18L L36P A59V | G28A T79A G150A C441T T443C C579G G633A T683C C852T G955A G968T G979A A1081G C1106T C1217T T1308C | T10A M27L – – A148V – – T228M – K319E I323S K327E A361T V369A V406A – | C145T A406G T541G T543C A545G C558T G662A A760G G775A C914T C944T G1019A T1082C G1146A | H49Y K136E C181G C181G Q182R – R221K M254V G259R A305V A315V R340H V361A – | C7T T21C C253T A293G G307A T389C T489C T498G A540G T581C A699G A890G C1014T A1047G A1067G T1085C G1114A T1176C C1249T T1366A G1489A G1491A A1493T G1512A A1580T C1587T A1619C G1622C A1623G T1625G T1626A C1627G C1628G | P3S – R85C K98R A103T M130T – – – L194S – Y297C – – Q356R L362P A372T – R417C W456R A497T A497T E498V – E527V – N540T R541P R541P I542R I542R P543G P543G |
Σ Н: 4 | Σ А: 4 | Σ Н: 16 | Σ А: 10 | Σ Н: 14 | Σ А: 11 | Σ Н: 33 | Σ А: 19 |
Note. N – nucleotide substitution; A – amino acid substitution. The position of a nucleotide or amino acid substitution is shown from the beginning of a gene.
The analysis of the nucleotide and amino acid substitutions showed that they, to a greater degree, differed from the known substitutions typical of the studied region [7]. Such significant differences can be explained by the fact that most of the CHIKV isolates were obtained by other researchers from sera of infected individuals and only rarely from mosquitoes. In addition, most of the isolates were obtained from mosquitoes captured in urban areas and rarely – in remote rural areas like mosquitoes in this study. The differences in nucleotide sequences of CHIKV isolates in all the 4 genes can also be explained by differences in locations of captured mosquitoes and by different mosquito species that carried the detected virus.
Thus, CHIKV and dengue type 1 and type 2 viruses were detected when isolating viruses from Aedes spp., Culex spp. and Culiseta spp. mosquitoes captured in forests in Nicaragua. The analysis of some CHIKV genes demonstrated high genetic diversity among viruses isolated from A. albopictus and A. aegypti mosquitoes.
The phylogenetic analysis of the nucleotide sequences (Fig. 2) showed that the closest relative of the isolated CHIKV is the virus isolated from a human in Africa in 2011 (GenBank KJ679577). The emergence of the common ancestor of these strains is estimated to have occurred 21 years ago (95% HPD 10–35). The isolate under study is also included in the cluster with prototype strains isolated in 1952–1986 in Tanzania and Africa.

Fig. 2. Phylogenetic tree of CHIKV E2, 6K, E1 genome fragments. The nodes of the tree indicate the posterior probability > 0.75 and the age of the group. The virus under study is highlighted. The scale shows the branch length representing years.
1 Tamura K., Stecher G., Kumar S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021; 38(7):3022–3027. DOI: 10.1093/molbev/msab120
2 Drummond A.J., Rambaut A., Shapiro B., Pybus O.G. Bayesian coalescent inference of past population dynamics from molecular sequences. Mol. Biol. Evol. 2005;22(5):1185–1192.
DOI: 10.1093/molbev/msi103
3 Hasegawa M., Kishino H., Yano T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985;22(2):160–174. DOI: 10.1007/BF02101694
About the authors
Georgy M. Ignatyev
I. Mechnikov Research Institute for Vaccines and Sera
Email: oksanich@yahoo.com
ORCID iD: 0000-0002-9731-3681
D. Sci. (Med.), Professor, main expert, Laboratory of molecular biotechnology, I.I. Mechnikov Research Institute for Vaccines and Sera
Russian Federation, MoscowAlexey S. Oksanich
I. Mechnikov Research Institute for Vaccines and Sera
Author for correspondence.
Email: oksanich@yahoo.com
ORCID iD: 0000-0002-8600-7347
Cand. Sci. (Biol.), leading researcher, Laboratory of molecular biotechnology, I.I. Mechnikov Research Institute for Vaccines and Sera
Russian Federation, MoscowElena V. Kazakova
St. Petersburg Research Institute for Vaccines and Sera
Email: oksanich@yahoo.com
ORCID iD: 0000-0002-0218-6641
Deputy director for HR and organizational design, St. Petersburg Research Institute for Vaccines and Sera
Russian Federation, St. PetersburgTatyana G. Samartseva
I. Mechnikov Research Institute for Vaccines and Sera
Email: oksanich@yahoo.com
ORCID iD: 0000-0003-3264-6722
researcher, Laboratory of molecular biotechnology, I.I. Mechnikov Research Institute for Vaccines and Sera
Russian Federation, MoscowЕlena V. Otrashevskaya
St. Petersburg Research Institute for Vaccines and Sera
Email: oksanich@yahoo.com
ORCID iD: 0000-0002-2491-4072
researcher, Laboratory of molecular biotechnology, I.I. Mechnikov Research Institute for Vaccines and Sera
Russian Federation, St. PetersburgStanislav V. Uyba
Mechnikov Latin American Institute of Biotechnology
Email: oksanich@yahoo.com
ORCID iD: 0000-0001-9246-3915
General director, Mechnikov Latin American Institute of Biotechnology
Nicaragua, ManaguaVictor P. Trukhin
St. Petersburg Research Institute for Vaccines and Sera
Email: oksanich@yahoo.com
ORCID iD: 0000-0002-6635-363X
Director, St. Petersburg Research Institute for Vaccines and Sera
Russian Federation, St. PetersburgReferences
- Kraemer M.U.D., Sinka M.E., Duda K.A., et al. The global compendium of Aedes aegypti and Aedes albopictus occurrence. Sci. Data. 2015;2:150035. DOI: http://doi.org/10.1038/sdata.2015.35
- Ponce P., Morales D., Argoti A., Cevallos V.E. First report of Aedes (Stegomyia) albopictus (Skuse) (Diptera: Culicidae), the Asian tiger mosquito, in Ecuador. J. Med. Entomol. 2018;55(1):248–9. DOI: http://doi.org/10.1093/jme/tjx165
- Hermanns K., Marklewitz M., Zirkel F., et al. Agua Salud alphavirus defines a novel lineage of insect-specific alphaviruses discovered in the New World. J. Gen. Virol. 2020;101(1):96–104. DOI: https://doi.org/10.1099/jgv.0.001344
- Guzman M.G., Halstead S.B., Artsob H., et al. Dengue: a continuing global threat. Nat. Rev. Microbiol. 2010;8(12 Suppl.):S7–16. DOI: http://doi.org/10.1038/nrmicro2460
- Vega-Rua A., Zouache K., Caro V., et al. High efficiency of temperate Aedes albopictus to transmit Chikungunya and dengue viruses in the Southeast of France. PLoS One. 2013;8(3):e59716. DOI: http://doi.org/10.1371/journal.pone.0059716
- Lundstrom J.O., Hesson J.C., Schafer M.L., et al. Sindbis virus polyarthritis outbreak signalled by virus prevalence in the mosquito vectors. PLoS Negl. Trop. Dis. 2019;13(8):e0007702. DOI: https://doi.org/10.1371/journal.pntd.0007702
- Villero-Wolf Y., Mattar S., Puerta-González A., et al. Genomic epidemiology of Chikungunya virus in Colombia reveals genetic variability of strains and multiple geographic introductions in outbreak, 2014. Sci. Rep. 2019;9(1):9970. DOI: https://doi.org/10.1038/s41598-019-45981-8
- Cevallos V., Ponce P., Waggoner J.J., et al. Zika and Chikungunya virus detection in naturally infected Aedes aegypti in Ecuador. Acta Trop. 2018;177:74–80. DOI: http://doi.org/10.1016/j.actatropica.2017.09.029
- Waggoner J.J., Gresh L., Vargas M.J., et al. Viremia and clinical presentation in Nicaraguan patients infected with Zika virus, Chikungunya virus, and Dengue virus. Clin. Infect. Dis. 2016;63(12):1584–90. DOI: http://doi.org/10.1093/cid/ciw589
- da Costa C.F., da Silva A.V., do Nascimento V.A., et al. Evidence of vertical transmission of Zika virus in field-collected eggs of Aedes aegypti in the Brazilian Amazon. PLoS Negl. Trop. Dis. 2018;12(7):e0006594. DOI: https://doi.org/10.1371/journal.pntd.0006594
- Eldadah Z.A., Asher D.M., Godec M.S., et al. Detection of flaviviruses by reverse-transcriptase polymerase chain reaction. J. Med. Virol. 1991;33(4):260–7. DOI: https://doi.org/10.1002/jmv.1890330410
- Игнатьев Г.М., Каа К.В., Оксанич А.С. и др. Индикация и идентификация вирусов денге и Чикунгунья в комарах рода Aedes spp., отловленных в центральной Америке. Журнал микробиологии, эпидемиологии и иммунобиологии. 2020;97(3):227–32. Ignatyev G.M., Kaa K.V., Oksanich A.S., et al. Indication and identification of dengue and Chikungunya viruses in Aedes spp. Mosquitoes captured in Central America. Journal of Microbiology, Epidemiology and Immunobiology. 2020;97(3):227–32 DOI: https://doi.org/10.36233/0372-9311-2020-97-3-4 EDN: https://elibrary.ru/ufhtab
- Čabanová V., Tichá E., Bradbury R.S., et al. Mosquito surveillance of West Nile and Usutu viruses in four territorial units of Slovakia and description of a confirmed autochthonous human case of West Nile fever, 2018 to 2019. Euro Surveill. 2021;26(19):2000063. DOI: https://doi.org/10.2807/1560-7917.S.2021.26.19.2000063
- Отрашевская Е.В., Казакова Е.В., Жиренкина Е.Н. и др. Ретроспективный серологический анализ распространения флавивирусных лихорадок и лихорадки Чикунгунья в Никарагуа; авидность специфических антител, как инструмент дифференциальной диагностики. Журнал микробиологии, эпидемиологии и иммунобиологии. 2022;99(2):215–24. Atrasheuskaya A.V., Kazakova E.V., Zhirenkina E.N., et al. The study of flaviviruses and Chikungunya virus seroprevalence in Nicaragua – virus-specific antibody avidity assay as a tool for differential diagnosis. Journal of Microbiology, Epidemiology and Immunobiology. 2022;99(2):215–24. DOI: https://doi.org/10.36233/0372-9311-196 EDN: https://elibrary.ru/tyopaj
- Shurtleff A.C., Beasley D.W., Chen J.J., et al. Genetic variation in the 3′ non-coding region of dengue viruses. Virology. 2001;281(1):75–87. DOI: https://doi.org/10.1006/viro.2000.0748
Supplementary files

