


Production of a new variant of soluble trimer Env of HIV-1 CRF63_02A6 SOSIP.664
- Authors: Rudometova N.B.1, Rudometov A.P.1, Fando A.A.1, Vahitov D.I.1, Ilyichev A.A.1, Agafonov A.P.1, Karpenko L.I.1
-
Affiliations:
- State Research Center of Virology and Biotechnology «Vector»
- Issue: Vol 101, No 5 (2024)
- Pages: 641-649
- Section: ORIGINAL RESEARCHES
- URL: https://microbiol.crie.ru/jour/article/view/18684
- DOI: https://doi.org/10.36233/0372-9311-590
- EDN: https://elibrary.ru/qxiziz
- ID: 18684

Cite item

Abstract
Introduction. Obtaining stabilized recombinant HIV-1 Env trimers that have a close to the native conformation is one of the directions in the field of development of vaccines against HIV-1.
The aim of the study was to obtain and characterize the stabilized trimer Env of HIV-1 SOSIP.664 based on the circulating genetic variant of the recombinant form CRF63_02A6.
Materials and methods. Bioinformatics resources were used to design the trimer Env gene based on the HIV-1 recombinant genetic variant CRF63_02A6. The designed gene was synthesized and cloned as part of an integration plasmid vector. A stable producer of trimer Env was obtained by transfection of the CHO-K1 cell line using the developed plasmid vector. Purification of the protein complex was performed using affinity chromatography and gel filtration. Antigenic properties of trimer Env were studied using immunochemical analysis using broadly neutralizing HIV-1 monoclonal antibodies (bnAbs).
Results. A new variant of the stabilized trimer Env of the surface glycoprotein of the recombinant form CRF63_02A6 HIV-1 SOSIP.664 was designed, including additional stabilizing modifications. Based on the CHO-K1 cell line, a stable producer was obtained and a purification protocol for the designed trimer Env was developed. It was found that the trimer Env CRF63_02A6 SOSIP.664 is effectively recognized by bnAbs 2G12, VRC01 and PGT126.
Conclusion. The obtained results indicate the prospects for further study of the structural features of the trimer Env CRF63_02A6 SOSIP.664, as well as its immunogenicity and the possibility of using it as a vaccine antigen.
Keywords
Full Text
Introduction
According to the Joint United Nations Program on HIV/AIDS (UNAIDS), the total number of people worldwide living with HIV-1 in 2023 was 39.9 million, and the number of recent HIV infections was more than 1 million1. In Russia during the year 2023, there were about 1.2 million people with a confirmed diagnosis of HIV infection2. The development of an effective vaccine to prevent HIV infection continues to be one of the most pressing unresolved problems.
Induction of an immune response capable of preventing infection with different HIV-1 subtypes is a critical goal for vaccines with the purpose of protection against HIV-1 infection. It is known that broadly neutralizing HIV-1 antibodies (bnAbs) can prevent HIV-1 infection by binding to envelope glycoproteins (Env) on the virion surface [1–4]. Work is actively underway to obtain native Env complexes to study their immunogenicity and determine their suitability as a vaccine antigen [5–8]. Over the last decade, significant progress has been made in this field. For example, various modifications, including SOSIP.664, have been developed to obtain a stabilized structure of recombinant Env trimers close to the native conformation and thus increase their ability to induce desired NAb responses against heterologous viruses [6, 9, 10]. Variants of stabilized Env trimers of genetic subtypes of HIV-1 (A, B, and C) circulating predominantly in Europe, America, and Africa were obtained, which showed the ability to induce neutralizing antibodies [6, 8, 11].
The recombinant form CRF63_02A6 is currently dominant in Siberian regions [12, 13]. According to M.V. Sivay et al., the recombinant form CRF63_02A6 was first registered in the Novosibirsk region in 2006. [14]. After that, it actively spread to other territories of Siberia (Omsk, Altai, Tomsk, Krasnoyarsk, and Kemerovo), displacing the A6 subtype and becoming the dominant strain. By 2020, CRF63_02A6 was detected in 20 regions of Russia and 6 countries of Central Asia. A study by the authors showed that CRF63_02A6 is detected in more than 80% of new HIV-1 cases in Siberia [14].
The aim of this study was to obtain a stabilized trimer Env recombinant form CRF63_02A6 of HIV-1 SOSIP.664, including additional stabilizing modifications.
Materials and methods
Monoclonal antibodies, bacterial strains, cell lines
Monoclonal antibodies PGT126, 2G12, VRC01 were obtained from the NIH HIV Reagent Program. CHO-K1 cell line was taken from the Collection of Cell Cultures of State Research Center of Virology and Biotechnology "Vector".
Design of an Env stabilized trimer of the recombinant form CRF63_02A6
The nucleotide sequence encoding the HIV-1 env trimer was designed based on the env 22RUAR13 gene of the recombinant form CRF63_02A6 (Collection of bacteria, bacteriophages and fungi of FBUNC Vector, Rospotrebnadzor, registration number P-124) using the SnapGene v. 3.2.1 and BioEdit v. 7.2.5 programs. Simulations were performed using Alphafold2 neural network colab program3; visualized using RCSB PDB website online tool, Mol*Plagin 3.43.1 3D Viewer tool. After the design process, codon optimization of the gene for expression in mammalian cells was carried out using online Codon Adaptation Tool4. The synthesis of the gene was carried out by DNA Synthesis. The obtained gene was cloned as part of an integration plasmid vector5. The integrity of the genetic construct was confirmed by sequencing (SB RAS Genomics Core Facility of the Institute of Chemical Biology and Fundamental Medicine).
Obtaining a stable producer of the trimer Env recombinant form CRF63_02A6 SOSIP.664
A stable Env trimer producer was obtained from the CHO-K1 cell line as described in [15]. After obtaining a polyclonal cell pool, it was used to isolate monoclonal cell cultures. For this purpose, the polyclonal cell culture was seeded in a 96-well plate at 1 cell per well in 200 μl of complete DMEM/F12 growth medium (Servicebio) containing 10% fetal bovine serum (HiMedia) and the antibiotic puromycin (Invivogen) at a concentration of 10 μg/mL and grown until a monolayer was achieved within 7-14 days. After this time, the grown monoclonal cultures were crossed into a 24-well plate. After the monoclonal cell lines formed a monolayer, they were analyzed for transgene expression using dot-blot analysis.
The selected monoclonal culture was then used for protein production. The monoclonal cell culture CHO-K1-gp140.SOSIP.664.opt was cultured in roller vials for 7 days, then the culture medium was collected and centrifuged to remove cellular debris. A purification procedure was performed using IMAC Seplife FF (Sunresin). The degree of purification of the target protein was assessed by polyacrylamide gel electrophoresis (PAAG) under denaturing conditions in the presence/absence of reducing agents followed by Coomassie G250 staining. Fractions containing the target protein were pooled and dialyzed against buffer (75 mM NaCl, 10 mM Tris, pH 8.0); concentrated using a 100 kDa centrifuge concentrator (Jet Biofil). Affinity-purified proteins were then separated by exclusion chromatography using a Chrom-LinXTM 16/1000 Tiderose GF200 column (Taidu Biotech) at a flow rate of 1 min/mL.
Dot-blot and western blot analyses
Dot-blot and Western blot analyses were performed according to standard methods using the SNAP i.d. 2.0 system (Millipore) and bnAb PGT126. Goat antibodies against human IgG conjugated with alkaline phosphatase (Sigma) were used as secondary antibodies. The immune complex was visualized by adding NBT/BCIP solution (Sigma).
Enzyme immunoassay
Enzyme immunoassay was performed according to standard methods. The primary antibodies used were bnAbs PGT126, 2G12, VRC01, and secondary antibodies were horseradish peroxidase-labeled goat anti-human IgG antibodies (Sigma). Tetramethylbenzidine solution (Imtek) was used as a chromogenic substrate. The reaction was stopped with 1 N hydrochloric acid solution. The optical density was measured on a Varioskan LUX instrument (Thermo Scientific) at a wavelength of 450 nm.
Statistical processing of data was performed using the GraphPad Prism 9 program (GraphPad Software Inc.).
Results
In this study, the characterized env 22RUAR13 gene, which belongs to the current recombinant form CRF63_02A6 of HIV-1 currently circulating in the Siberian region, was used for the design. To improve the formation of soluble trimers, the construct contained the following modifications compared to the natural sequence: the natural signal peptide was replaced by the tPA (tissue plasminogen activator signal peptide) signal sequence; 2 cysteines were added to form the gp120-gp41 disulfide bond (501C and 605C); added stabilizing amino acid substitutions in gp41 (I559P, H66R and A316W); added amino acid substitutions in gp41 (I535M and Q543N) and TD8 mutation complex (E47D, N49E, V65K, E106T, I165L, G429R and K500R; 432Q); to enhance cleavage, the REKR motif in gp120 was replaced with RRRRRR (R6); the MPER region, transmembrane and cytoplasmic domains were also deleted for better expression of the designed protein; after amino acid residue 664 at the C-terminus, 6×His was added for further purification [6] (Fig. 1). The resulting Env trimer variant of the recombinant form CRF63_02A6 was designated as gp140.SOSIP.664.opt.

Fig. 1. Design of the Env trimer gp140.SOSIP.664.opt.
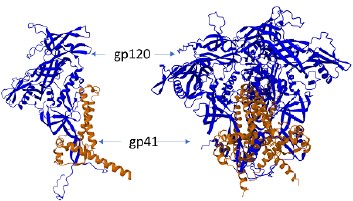
Spatial structure modeling showed that the designed amino acid sequence was capable of forming specific monomers and trimeric complexes (Fig. 2).

Fig. 2. Structure model of the monomer (а) and trimer (b) of gp140.SOSIP.664.opt.
Furthermore, a polyclonal cell culture CHO-K1-gp140.SOSIP.664.opt was obtained by transfection of the CHO-K1 cell line, which was used to isolate monoclonal cell lines by the limiting dilution method. As a result, 24 monoclonal cultures were obtained and examined for their ability to produce the target protein into the culture medium using dot-blot analysis. As a result, a monoclonal culture (No. 11) with high transgene expression was identified (Fig. 3), which was used for further research.

Fig. 3. Dot blot analysis of the culture medium collected from monoclonal cell cultures CHO-K1-gp140.SOSIP.664.opt.
Recombinant Env trimer CRF63_02A6 was used as C+, and culture medium collected from non-transformed CHO-K1 cells was used as C–.
For preparative production of the target protein, monoclonal cell culture of CHO-K1-gp140.SOSIP.664.opt was cultured in roller bottles followed by purification of the recombinant Env trimer. At the end of cultivation, the culture medium was collected, and the target protein was purified by metal-chelate affinity chromatography, dialysis and subsequent gel filtration. The protein production was about 15 mg/L.
The purified Env protein product was characterized by PAGE electrophoresis in the presence or absence of β-mercaptoethanol. Based on the amino acid sequence of recombinant Env CRF63_02A6 (683 a. a.), the theoretically calculated molecular mass of the monomer should be 76.8 kDa. However, it is known that in eukaryotic cells the Env HIV-1 protein is highly glycosylated and about 50% of its molecular mass can be made up of glycans [16, 17], resulting in an increase in the mass of the monomer to 140 kDa. In the eukaryotic cell culture, glycosylation of Env protein may be incomplete and there may be glycosylation intermediates. For this reason, in the case of electrophoresis under denaturing conditions (in the presence of β-mercaptoethanol), 2 bands in PAGE are observed (Fig. 4, a), differing by approximately 10-20 kDa, which represent monomers of the recombinant Env trimer with different degrees of glycosylation. Both are recognized by the monoclonal antibody PGT126 (Fig. 4, b).

Fig. 4. Characterization of the Env protein.
a — electropherogram of separation of recombinant Env trimer gp140.SOSIP.664.opt in 12% PAGE: 1 — molecular weight marker of proteins Precision Plus Dual Color Standards (Bio-Rad, USA); 2 — in the presence of β-mercaptoethanol; 3 — in the absence of β-mercaptoethanol. b — immunoblotting of Env trimer gp140.SOSIP.664.opt with monoclonal antibody PGT126: 1 — molecular weight marker of proteins Precision Plus Dual Color Standards (Bio-Rad, USA); 2 — in the presence of β-mercaptoethanol; 3 — in the absence of β-mercaptoethanol; c — profile of size-exclusion chromatography of Env trimer gp140.SOSIP.664.opt on a Chrom-LinXTM 16/1000 Tiderose GF200 column.
In the absence of β-mercaptoethanol (Fig. 4, a, band 3), monomeric forms of protein are absent in PAGE, and higher order oligomers (more than 250 kDa) are detected.
Using gel filtration on a pre-calibrated ChromLinXTM 16/1000 Tiderose GF200 column (Taidu Biotech), it was found that the main peak of the target protein comes out in the region corresponding to the trimeric form (Fig. 4, c).
To study the antigenic properties of the Env trimer, an enzyme immunoassay was performed using bnAbs 2G12, VRC01 and PGT126 targeting an epitope including carbohydrate residues at positions 295, 332 and 392, a CD4-binding site and a V3-loop on the surface of gp120, respectively. The resulting Env gp140.SOSIP.664 trimer.opt was found to be efficiently recognized by monoclonal broadly neutralizing antibodies (Fig. 5), which may indicate that the Env trimer synthesized in the producer cells is properly stacked and maintains its conformation. The EC50 values for bnAbs 2G12, VRC01 and PGT126 were 0.313, 0.098 and 0.019 μg/mL, respectively (Table).

Fig. 5. Specific activity of monoclonal broadly neutralizing antibodies against Env trimer of the recombinant form CRF63_02A6 gp140.SOSIP.664.opt (presented as a heat map). The amount of absorbed antigen was 100 ng/well.
EC50 values calculated for gp140.SOSIP.664.opt compared with literature data
bnAbs | EC50 μg/mL | |||
gp140.SOSIP.664.opt (CRF63_02A6) | BG505 (А) | JR-FL (B) | B41 (B) | |
PGT126 | 0,019 | N. d. | N. d. | |
2G12 | 0,313 | 0,21 [19] | 0,13 [19] | |
VRC01 | 0,098 | 0,67 [19] | 5,20 [19] | |
Note. N. d. — no data.
Discussion
Env trimers of various HIV-1 subtypes have been actively investigated worldwide in terms of both structural and antigenic features and immunogenic properties [6–8, 11]. Thus, native Env trimers of clades A, B, C, and G [20–26], as well as Env trimers based on the consensus sequence of clade C [27] and group M [6], respectively, have been characterized in detail using biochemical and structural approaches.
Predominantly, studies have focused on subtypes A, B, and C, with virtually no data on various CRFs [28], in particular the recombinant form CRF63_02A6, which is rapidly spreading in Russian regions and is currently dominant in the Siberian Federal District [12–14].
We previously obtained an Env trimer of the recombinant form CRF63_02A6 HIV-1, the nucleotide sequence of the env gene of the recombinant form CRF63_02A6 HIV-1 and known modifications of SOSIP.664 were also used in the design of this trimer [29]: Intermolecular disulfide bond (SOS, A501C-T605C) for binding gp120 and gp41; improved furin cleavage site (RRRRRR); point substitution of I559P, which helps stabilize gp41 subunits in a pre-fusion conformation; deletion of MPER region, transmembrane and cytoplasmic domains for better expression; addition of 6×His at the C-terminus for subsequent purification; the signal peptide was left natural. Using immunoassays, it is shown that CHO-K1 trimers synthesized in cells are recognized by both monoclonal broadly neutralizing antibodies and sera from HIV-positive patients [15].
Later, high-resolution crystal structures of BG505 SOSIP were obtained, which made it possible to identify additional positions of amino acid residues involved in trimer stabilization and contributing to the formation of well-ordered, homogeneous, and highly stable soluble trimers [6, 30]. The modified BG505 SOSIP.664-140 Env trimers obtained taking into account these data demonstrated the ability to induce a specific humoral immune response in laboratory animals. It was shown that such SOSIP.664-140 complex is actively recognized by neutralizing monoclonal antibodies, and, on the contrary, non-neutralizing antibodies do not bind to it [18].
In this study, bioinformatic resources were used to design an Env trimer gp140.SOSIP.664.opt of recombinant form CRF63_02A6, in the structure of which, along with the SOS disulfide bond (A501C-T605C) and the amino acid substitution I559P, we introduced additional amino acid substitutions and the TD8 mutation complex in gp120, leading to additional stabilization of the protein complex [6]. As a result, an optimized stabilized trimer of Env CRF63_02A6 HIV-1 was obtained. Computer modeling using Alphafold2 showed that the designed amino acid sequence is able to fold into monomers and trimeric complexes (Fig. 1, b).
Using PAGE electrophoresis and Western blot analysis, we found that in the presence of β-mercaptoethanol, the purified HIV-1 Env trimer CRF63_02A6 denatures to a monomer (molecular mass ~140 kDa; Fig. 4, a) that is specifically recognized by bnAb PGT126 (Fig. 4, b). In the absence of β-mercaptoethanol, the monomer fraction is absent, indicating that the protein tends to aggregate into larger structures (Fig. 4, a). Gel filtration revealed that the protein in solution is predominantly in trimeric form (Fig. 4, c).
When CHO-K1-gp140.SOSIP.664.opt cells were cultured, the Env trimer was detected in the culture medium, indicating that it is soluble. The fact that the obtained Env trimer is efficiently recognized by monoclonal broadly neutralizing antibodies bnAbs 2G12, VRC01 and PGT126 (Fig. 5), which recognize conformational epitopes on the Env surface within the viral particle, testifies to its proper folding and preservation of conformation. It should be noted that the EC50 values obtained for the gp140.SOSIP.664.opt Env trimer differ from the EC50 values for Env trimers of other HIV-1 subtypes (Table). This may indicate the presence of antigenic differences between the above-mentioned HIV-1 strains.
Conclusion
An optimized stabilized trimer of Env HIV-1 gp140.SOSIP.664.opt based on the actual circulating genetic variant of the recombinant form CRF63_02A6 was created and characterized. The obtained data indicate the prospect of further study of its structural features, immunogenicity and the possibility of using it as a vaccine antigen.
1 UNAIDS Fact Sheet 2024. Global HIV statistics. 2024. URL: https://www.unaids.org/sites/default/files/media_asset/UNAIDS_FactSheet_ru.pdf (date of access: 28.09.2024).
2 HIV infection in the Russian Federation as of June 30, 2023. 2023. URL: https://files.antispidnn.ru/uploads/docs/spec/vich2023.pdf (date of reference: 18.09.2024).
3 URL: https://colab.research.google.com/github/sokrypton/ColabFold/blob/main/AlphaFold2.ipynb
4 URL: https://www.jcat.de
5 Patent No. 2800471 Russian Federation, MPK C12N15/00 (2006.01), C07K16/10 (2006.01), C12N1/00 (2006.01), C12N15/63 (2006.01). Plasmid genetic construct pVEAL3-10H10ch, recombinant cell line strain CHO-K1-10H10ch and chimeric antibody 10H10ch against tick-borne encephalitis virus produced by said cell line strain CHO-K1-10H10ch: 2022126714: applied. 13.10.2022: publ. 21.07.2023 / Shanshin D.V., Nesmeyanova V.S., Shcherbakov D.N. et al. URL: https://patents.google.com/patent/RU2800471C1/ru (date of reference: 18.09.2024).
About the authors
Nadezhda B. Rudometova
State Research Center of Virology and Biotechnology «Vector»
Author for correspondence.
Email: nadenkaand100@mail.ru
ORCID iD: 0000-0002-1684-9071
Cand. Sci. (Biol.), senior researcher, Bioengineering department
Russian Federation, KoltsovoAndrey P. Rudometov
State Research Center of Virology and Biotechnology «Vector»
Email: nadenkaand100@mail.ru
ORCID iD: 0000-0003-2808-4309
Cand. Sci. (Biol.), leading researcher, Bioengineering department
Russian Federation, KoltsovoAnastasia A. Fando
State Research Center of Virology and Biotechnology «Vector»
Email: nadenkaand100@mail.ru
ORCID iD: 0000-0002-3325-3325
research intern, Bioengineering department
Russian Federation, KoltsovoDanil I. Vahitov
State Research Center of Virology and Biotechnology «Vector»
Email: nadenkaand100@mail.ru
ORCID iD: 0009-0008-0706-8119
graduate student, research intern, Bioengineering department
Russian Federation, KoltsovoAlexander A. Ilyichev
State Research Center of Virology and Biotechnology «Vector»
Email: nadenkaand100@mail.ru
ORCID iD: 0000-0001-5356-0843
D. Sci. (Biol.), Professor, Head, Bioengineering department
Russian Federation, KoltsovoAlexander P. Agafonov
State Research Center of Virology and Biotechnology «Vector»
Email: nadenkaand100@mail.ru
ORCID iD: 0000-0003-2577-0434
D. Sci. (Biol.), General director, State Research Center of Virology and Biotechnology "Vector
Russian Federation, KoltsovoLarisa I. Karpenko
State Research Center of Virology and Biotechnology «Vector»
Email: nadenkaand100@mail.ru
ORCID iD: 0000-0003-4365-8809
D. Sci. (Biol.), leading researcher, Bioengineering department
Russian Federation, KoltsovoReferences
- Jones L.D., Moody M.A., Thompson A.B. Innovations in HIV-1 vaccine design. Clin. Ther. 2020;42(3):499–514. DOI: https://doi.org/10.1016/j.clinthera.2020.01.009
- Kumar S., Singh S., Luthra K. An overview of human anti-HIV-1 neutralizing antibodies against diverse epitopes of HIV-1. ACS Omega. 2023;8(8):7252–61. DOI: https://doi.org/10.1021/acsomega.2c07933
- Thavarajah J.J., Hønge B.L., Wejse C.M. The use of broadly neutralizing antibodies (bNAbs) in HIV-1 treatment and prevention. Viruses. 2024;16(6):911. DOI: https://doi.org/10.3390/v16060911
- Caskey M. Broadly neutralizing antibodies for the treatment and prevention of HIV infection. Curr. Opin. HIV AIDS. 2020;15(1):49–55. DOI: https://doi.org/10.1097/COH.0000000000000600
- Vzorov A.N., Wang L., Chen J., at al. Effects of modification of the HIV-1 Env cytoplasmic tail on immunogenicity of VLP vaccines. Virology. 2016;489:141–50. DOI: https://doi.org/10.1016/j.virol.2015.09.015
- Sliepen K., Han B., Bontjer I., et al. Structure and immunogenicity of a stabilized HIV-1 envelope trimer based on a group-M consensus sequence. Nat. Commun. 2019;10(1):2355. DOI: https://doi.org/10.1038/s41467-019-10262-5
- Olia A.S., Cheng C., Zhou T., et al. Soluble prefusion-closed HIV-envelope trimers with glycan-covered bases. iScience. 2023;26(8):107403. DOI: https://doi.org/10.1016/j.isci.2023.107403
- Zhao F., Joyce C., Burns A., et al. Mapping neutralizing antibody epitope specificities to an HIV Env trimer in immunized and in infected rhesus macaques. Cell Rep. 2020;32(10):108122. DOI: https://doi.org/10.1016/j.celrep.2020.108122
- Sanders R.W., Moore J.P. Native-like Env trimers as a platform for HIV-1 vaccine design. Immunol. Rev. 2017;275(1):161–82. DOI: https://doi.org/10.1111/imr.12481
- Torrents de la Peña A., Sanders R.W. Stabilizing HIV-1 envelope glycoprotein trimers to induce neutralizing antibodies. Retrovirology. 2018;15(1):63. DOI: https://doi.org/10.1186/s12977-018-0445-y
- Schorcht A., van den Kerkhof T.L.G.M., Cottrell C.A., et al. Neutralizing antibody responses induced by HIV-1 envelope glycoprotein SOSIP trimers derived from elite neutralizers. J. Virol. 2020;94(24):e01214-20. DOI: https://doi.org/10.1128/jvi.01214-20.
- Maksimenko L.V., Totmenin A.V., Gashnikova M.P., et al. Genetic diversity of HIV-1 in Krasnoyarsk Krai: Area with high levels of HIV-1 recombination in Russia. Biomed. Res. Int. 2020; 2020:9057541. DOI: https://doi.org/10.1155/2020/9057541
- Rudometova N.B., Shcherbakova N.S., Shcherbakov D.N., et al. Genetic diversity and drug resistance mutations in reverse transcriptase and protease genes of HIV-1 isolates from Southwestern Siberia. AIDS Res. Hum. Retroviruses. 2021;37(9):716–23. DOI: https://doi.org/10.1089/AID.2020.0225
- Sivay M.V., Maksimenko L.V., Osipova I.P., et al. Spatiotemporal dynamics of HIV-1 CRF63_02A6 sub-epidemic. Front. Microbiol. 2022;13:946787. DOI: https://doi.org/10.3389/fmicb.2022.946787
- Rudometova N.B., Rudometov A.P., Fando A.A., et al. Production and study of immunochemical properties of stabilized Env trimer of recombinant form CRF63_02A6 of HIV-1. Bull. Exp. Biol. Med. 2023;176(1):96–100. DOI: https://doi.org/10.1007/s10517-023-05978-w
- Rathore U., Saha P., Kesavardhana S., et al. Glycosylation of the core of the HIV-1 envelope subunit protein gp120 is not required for native trimer formation or viral infectivity. J. Biol. Chem. 2017;292(24):10197–219. DOI: https://doi.org/10.1074/jbc.M117.788919
- Wang W., Nie J., Prochnow C., et al. A systematic study of the N-glycosylation sites of HIV-1 envelope protein on infectivity and antibody-mediated neutralization. Retrovirology. 2013;10:14. DOI: https://doi.org/doi.org/10.1186/1742-4690-10-14
- Sanders R.W., Derking R., Cupo A., et al. A next-generation cleaved, soluble HIV-1 Env trimer, BG505 SOSIP.664 gp140, expresses multiple epitopes for broadly neutralizing but not non-neutralizing antibodies. PLoS Pathog. 2013;9(9):e1003618. DOI: https://doi.org/10.1371/journal.ppat.1003618
- Cao L., Pauthner M., Andrabi R., et al. Differential processing of HIV envelope glycans on the virus and soluble recombinant trimer. Nat. Commun. 2018;9(1):3693. DOI: https://doi.org/10.1038/s41467-018-06121-4
- Julien J.P., Cupo A., Sok D., et al. Crystal structure of a soluble cleaved HIV-1 envelope trimer. Science. 2013;342(6165):1477–83. DOI: https://doi.org/10.1126/science.1245625
- Lyumkis D., Julien J.P., de Val N., et al. Cryo-EM structure of a fully glycosylated soluble cleaved HIV-1 envelope trimer. Science. 2013;342(6165):1484–90. DOI: https://doi.org/10.1126/science.1245627
- Pancera M., Zhou T., Druz A., et al. Structure and immune recognition of trimeric pre-fusion HIV-1 Env. Nature. 2014; 514(7523):455–61. DOI: https://doi.org/10.1038/nature13808.
- Garces F., Lee J.H., de Val N., et al. Affinity maturation of a potent family of HIV antibodies is primarily focused on accommodating or avoiding glycans. Immunity. 2015;43(6):1053–63. DOI: https://doi.org/10.1016/j.immuni.2015.11.007
- Lee J.H., Ozorowski G., Ward A.B. Cryo-EM structure of a native, fully glycosylated, cleaved HIV-1 envelope trimer. Science. 2016;351(6277):1043–8. DOI: https://doi.org/10.1126/science.aad2450
- Stewart-Jones G.B.E., Soto C., Lemmin T., et al. Trimeric HIV-1-Env structures define glycan shields from clades A, B, and G. Cell. 2016;165(4):813–26. DOI: https://doi.org/10.1016/j.cell.2016.04.010
- Guenaga J., Garces F., de Val N., et al. Glycine substitution at helix-to-coil transitions facilitates the structural determination of a stabilized subtype C HIV envelope glycoprotein Immunity. 2017;46(5):792–803. DOI: https://doi.org/10.1016/j.immuni.2017.04.014
- Rutten L., Lai Y.T., Blokland S., et al. A universal approach to optimize the folding and stability of prefusion-closed HIV-1 envelope trimers. Cell Rep. 2018;23(2):584–95. DOI: https://doi.org/10.1016/j.celrep.2018.03.061
- Niu J., Wang Q., Zhao W., et al. Structures and immune recognition of Env trimers from two Asia prevalent HIV-1 CRFs. Nat. Commun. 2023;14(1):4676. DOI: https://doi.org/10.1038/s41467-023-40321-x
- Pugach P., Ozorowski G., Cupo A., et al. A native-like SOSIP.664 trimer based on an HIV-1 subtype B env gene. J. Virol. 2015; 89(6):3380–95. DOI: https://doi.org/10.1128/JVI.03473-14
- Guenaga J., Dubrovskaya V., de Val N., et al. Structure-guided redesign increases the propensity of HIV Env to generate highly stable soluble trimers. J. Virol. 2015;90(6):2806–17. DOI: https://doi.org/10.1128/JVI.02652-15
Supplementary files

