


Детерминанты резистентности к левофлоксацину и метронидазолу российских клинических изолятов Helicobacter pylori по результатам полногеномного секвенирования
- Авторы: Старкова Д.А.1, Гладышев Н.С.2, Полев Д.Е.1, Саитова А.Т.1, Глазунова К.А.3, Егорова С.А.1, Сварваль А.В.1
-
Учреждения:
- Санкт-Петербургский научно-исследовательский институт эпидемиологии и микробиологии имени Пастера
- Российский научный центр хирургии имени академика Б.В. Петровского
- Санкт-Петербургский государственный университет
- Выпуск: Том 102, № 4 (2025)
- Страницы: 465-473
- Раздел: ОРИГИНАЛЬНЫЕ ИССЛЕДОВАНИЯ
- URL: https://microbiol.crie.ru/jour/article/view/18926
- DOI: https://doi.org/10.36233/0372-9311-710
- EDN: https://elibrary.ru/YYLGLL
- ID: 18926

Цитировать

Аннотация
Введение. Устойчивость к антибактериальным препаратам является одной из ключевых проблем в лечении Helicobacter pylori-инфекции, однако в России практически отсутствуют данные о механизмах резистентности к метронидазолу (МТZ) и левофлоксацину (LVX).
Цель работы — выявление детерминант резистентности у клинических изолятов H. pylori к МТZ и LVX с использованием данных полногеномного секвенирования.
Материалы и методы. Проведён ретроспективный анализ 43 изолятов H. pylori, выделенных от взрослых пациентов (2014–2022 гг.). Чувствительность к антибактериальным препаратам определяли бактериологическим диско-диффузионным методом. Полногеномное секвенирование 43 штаммов H. pylori проводили с использованием секвенатора «DNBSEQ-G50».
Результаты. Оценка результатов теста фенотипической лекарственной чувствительности показала, что 11 изолятов являлись чувствительными к МТZ (МТZ-S), 31 — чувствительными к LVX (LVX-S), тогда как 32 изолята проявляли устойчивость к МТZ (МТZ-R), 12 — к LVX (LVX-R). Для выявления ассоциации между фенотипической и генотипической устойчивостью проведён анализ нуклеотидных замен в генах gyrA, gyrB, rdxA, frxA, fdxB, fur. Из всех мутаций, выявленных в генах gyrA и gyrB, только D91/GNY в гене gyrA была ассоциирована с фенотипической устойчивостью к LVX и обнаружена у 4/12 (33,3%) изолятов (p < 0,05). Комбинированная мутация D91G/N/Y+N87K в гене gyrA выявлена у 6/12 (50,0%) LVX-R-изолятов (p < 0,001). У 21,9% (7/32) МТZ-R-изолятов в гене rdxA выявлены точечные мутации, приводящие к сдвигу рамки считывания или преждевременной терминации синтеза белка. Ни одна из мутаций в генах frxA, fur и fdxB не была ассоциирована с устойчивостью H. pylori к МТZ.
Выводы. По результатам полногеномного секвенирования российских клинических изолятов H. pylori детекция комбинированной мутации D91G/N/Y+N87K в гене gyrA может служить предиктором фенотипической устойчивости H. pylori к LVX.
Полный текст
Введение
Инфекция Helicobacter pylori остаётся одной из самых распространённых хронических бактериальных инфекций в мире и считается основным фактором риска развития рака желудка (около 90% случаев) [1]. Согласно ключевым положениям Маастрихт VI/Флорентийского консенсуса, лежащего в основе национальных клинических рекомендаций, инфекция H. pylori (независимо от симптомов или осложнений) всегда вызывает гастрит, единственным методом лечения которого является эрадикационная терапия. Кроме того, согласно первому положению, для рационального использования антибиотиков целесообразно проводить тесты на лекарственную чувствительность до назначения терапии 1-й линии [2, 3].
Устойчивость H. pylori к антибактериальным препаратам (АБП) признана на сегодняшний день одной из самых серьёзных проблем. Для эрадикации H. pylori используется комбинированная терапия, состоящая из 2–3 АБП: амоксициллин, кларитромицин, метронидазол (МТZ), тетрациклин, левофлоксацин (LVX), рифабутин; ингибитора протонной помпы и препаратов висмута (или без него) [2, 3]. Однако в последние годы наблюдается неуклонное снижение уровня эффективности антихеликобактерной терапии параллельно с развивающейся устойчивостью к противомикробным препаратам [4, 5]. Более того, широкое применение АБП или низкий уровень комплаенса при лечении приводят к развитию устойчивости не только H. pylori, но и других бактерий микробиома желудочно-кишечного тракта, способствуя селективному давлению АБП в сторону отбора генов резистентности, обогащению резистома и распространению среди бактериальных сообществ [6, 7].
Молекулярно-генетические механизмы, лежащие в основе развития антибиотикорезистентности (АБР) H. pylori, полностью не изучены. Принято считать, что устойчивость H. pylori к АБП обусловлена мутациями в хромосомной ДНК de novo, которые либо изменяют мишень действия АБП, либо препятствуют его активации внутри клеток [6]. Тем не менее у значительной доли штаммов H. pylori, устойчивых к АБП, отсутствуют известные генетические детерминанты резистентности. Это указывает на сложный и многофакторный характер механизмов устойчивости, выходящий за рамки мутационной активности и/или активации отдельных генов в ответ на применение АБП. К потенциальным механизмам устойчивости H. pylori относят:
- повышенную экспрессию генов системы эффлюкса;
- синергетические взаимодействия, включающие мутации, горизонтальный перенос генов и активацию защитных систем;
- клеточную адаптацию, связанную с образованием биопленок и коккоидов, устойчивых к АБП;
- компенсаторные мутации, нейтрализующие снижение фитнеса устойчивых штаммов за счёт эпистатического взаимодействия с детерминантами резистентности;
- гетерорезистентность — феномен, при котором в бактериальной популяции одновременно присутствуют субпопуляции клеток с разной чувствительностью к АБП.
Все перечисленные механизмы не только способствуют формированию АБР H. pylori, но и ускоряют развитие полирезистентности, что существенно усложняет процесс эрадикации и подчёркивает необходимость постоянного пересмотра и усовершенствования терапевтических подходов [6–8].
Механизм действия МТZ — препарата 1-й линии эрадикационной терапии H. pylori — заключается в восстановлении нитрогруппы в анаэробных условиях, что приводит к образованию цитотоксичных нитроанионов и свободных радикалов, повреждающих ДНК и нарушающих функционирование бактериальных клеток. Устойчивость H. pylori к МТZ обусловлена совокупностью сложных генетических и биохимических процессов, которые до настоящего времени полностью не изучены. Основной механизм резистентности к МТZ опосредован инактивацией генов ферментов редуктаз rdxA (кодирует нечувствительную к кислороду нитроредуктазу) и frxA (кодирует флавиноксидазу), что снижает способность МТZ восстанавливаться до активных форм (NO2– и NO22–) и, как следствие, уменьшает антимикробное действие препарата [9, 10]. Мутации в гене fur, регулирующем захват железа, и fdxB, кодирующем ферредоксин, предположительно также могут способствовать формированию АБР [7, 8, 10]. Повышение уровня антиоксидантных ферментов и системы эффлюкса, а также мутационной активности в генах, ответственных за репарацию повреждённой ДНК, выступают в роли дополнительных факторов, способствующих развитию резистентности H. pylori к МТZ [6].
Другой препарат, выступающий в качестве резервной терапии для лечения хеликобактериоза, — LVX — оказывает антибактериальный эффект за счёт ингибирования топоизомеразы II (ДНК-гиразы) и топоизомеразы IV — ключевых ферментов, участвующих в процессах репликации и рекомбинации ДНК. Наиболее распространённый механизм резистентности H. pylori к фторхинолонам обусловлен точечными мутациями в области QRDR (Quinolone Resistance-Determining Regions), определяющей резистентность к хинолонам: мутации в кодонах 86, 87, 88, 91, 97 гена gyrA и кодонах 481, 484 и 463 гена gyrB [6, 7, 10]. Однако роль некоторых из перечисленных мутаций в развитии устойчивости H. pylori к LVX не доказана.
В России практически отсутствуют систематизированные и достоверные данные о распространённости устойчивости H. pylori к LVX и МТZ. Кроме того, исследований, посвящённых выявлению генетических детерминант резистентности к данным АБП, практически нет, что затрудняет разработку эффективных молекулярно-генетических методов диагностики и мониторинга резистентности в клинической практике.
Целью нашего исследования явилось выявление детерминант резистентности у клинических изолятов H. pylori к МТZ и LVX с использованием данных полногеномного секвенирования.
Материалы и методы
Проведён ретроспективный анализ 43 клинических изолятов H. pylori, выделенных от взрослых пациентов с заболеваниями желудочно-кишечного тракта (2014–2022 гг.) в Санкт-Петербургском научно-исследовательском институте эпидемиологии и микробиологии им. Пастера. Средний возраст пациентов составлял 44,0 ± 4,5 года (22–70 лет).
Биоптаты слизистой оболочки антрального отдела и/или дна желудка, помещённые в тиоглеколевую среду, подлежали гомогенизации и последующему культивированию на селективной среде на основе Колумбийского агара с добавлением 7% дефибринированной лошадиной крови и 1% раствора IsoVitalex при 37°С в микроаэрофильных условиях (содержание кислорода ~ 5%) с использованием газогенерирующих пакетов GasPak 100 (BBL CampyPak Plus Microaerophilic System envelopes with Palladium Catalyst, «BD Biosciences»). Для идентификации H. pylori использовали набор биохимических тестов (уреазный, каталазный и оксидазный) и набор реагентов для выявления ДНК H. pylori методом полимеразной цепной реакции (ПЦР) («ДНК-Технология»).
Для постановки теста лекарственной чувствительности бактериальную культуру H. pylori суспендировали до плотности 2 по стандарту мутности Мак-Фарланда (~ 6 × 108 КОЕ/мл), наносили 0,1 мл на поверхность чашки Петри с агаром Мюллера–Хинтона с добавлением 5% дефибринированной лошадиной крови и равномерно распределяли по поверхности шпателем. Чувствительность изолятов H. pylori к МТZ и LVX определяли диско-диффузионным методом: сразу после инокуляции на поверхность агара асептически наносили диски с МТZ (5 мкг/диск) и LVX (5 мкг/диск) инкубировали в микроаэрофильных условиях (5% O2, 10% CO2, 85% N2) при 37°С в течение 72 ч. После окончания инкубации измеряли диаметр зон полного подавления роста вокруг диска с антибиотиком (мм). Интерпретацию результатов диско-диффузионного метода проводили на основании пороговых значений, представленных в публикации Z. Zhong и соавт.: штаммы H. pylori считались устойчивыми к МТZ (MTZ-R) при диаметре зоны ингибиции ≤ 16 мм, чувствительными (MTZ-S) при диаметре ≥ 17 мм; устойчивыми к LVX (LVX-R) — при диаметре зоны ингибиции ≤ 17 мм, чувствительными (LVX-S) — при диаметре ≥ 18 мм [11].
Хромосомную ДНК из чистых культур H. pylori выделяли с помощью набора «QIAamp DNA Mini Kit» («QIAGEN GmbH») в соответствии с инструкцией производителя. Концентрация ДНК каждого образца была количественно определена на флуориметре «Qubit 4.0». Полногеномное секвенирование проводили на секвенаторе «DNBSEQ-G50» («MGI Tech Co. Ltd.»).
Оценку качества парно-концевых библиотек, удаление адаптеров и последовательностей низкого качества (Q-score < 20) проводили с помощью программ «FastQC v. 0.12.1» и «Trim Galore! v. 0.6.7». Бактериальные геномы были собраны de novo с помощью геномного ассемблера «SPAdes v. 3.13.1», а результаты были оценены с помощью «QUAST v. 5.2.0» [12, 13]. Полученные геномные последовательности были выровнены на последовательность референсного штамма H. pylori 26695 (GenBank acc. no. AE000511.1). Для оценки генетических вариаций между изолятами и выявления нуклеотидных замен использовали программу «Snippy v. 4.6.0»1. Выровненные нуклеотидные последовательности были визуально проанализированы с помощью «UGENE v. 38.1» [14]. Все сборки геномов клинических изолятов H. pylori были депонированы в базу данных NCBI GenBank под регистрационным номером PRJNA10110372.
Статистическую обработку результатов исследования проводили с использованием языка программирования R v. 4.3.2. Наличие корреляции между двумя группами (фенотипической и генотипической) выявляли с использованием критерия χ2 и точного критерия Фишера. Различия между группами считали значимыми при p < 0,05.
Результаты
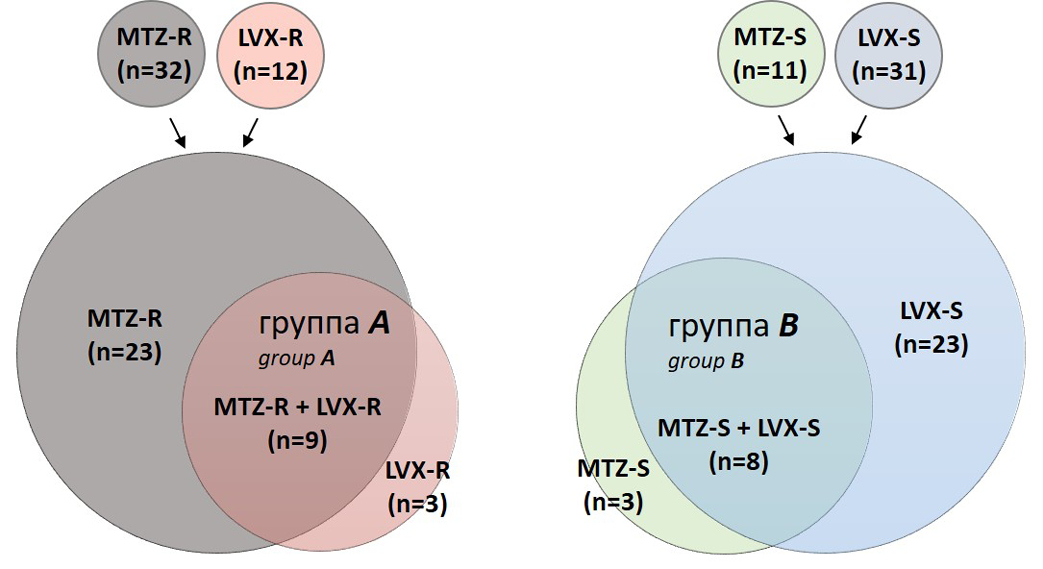
Анализ теста фенотипической лекарственной чувствительности показал, что среди 43 клинических изолятов H. pylori 12 проявляли устойчивость к LVX (LVX-R), 32 — к МТZ (MTZ-R); в то же время 31 изолят был чувствителен к LVX (LVX-S), 11 — к МТZ (MTZ-S). Среди резистентных изолятов 9 проявляли устойчивость одновременно к обоим препаратам (группа А), тогда как среди чувствительных изолятов 8 были чувствительны как к LVX, так и к МТZ (группа В: рисунок).

Диаграмма Венна, отображающая комбинации статусов фенотипической лекарственной чувствительности клинических изолятов H. pylori к MTZ и LVX.
Для поиска детерминант резистентности к LVX и MTZ и их ассоциации с фенотипической устойчивостью все 43 изолята были подвергнуты полногеномному секвенированию, после чего проведён анализ нуклеотидных замен в генах gyrA, gyrB, rdxA, frxA, fdxB и fur.
Из всех мутаций в гене gyrA только D91N/Y/G была ассоциирована с фенотипической лекарственной устойчивостью и выявлена у 33,3% (4/12) LVX-R-изолятов (p < 0,05) (таблица). Миссенс-мутация N87K в гене gyrA выявлена у 16,7% (2/12) LVX-R-изолятов и не встречалась в сочетании с мутациями D91N/Y/G и A88P, тогда как мутация A88P выявлена лишь у 1 (8,3%) устойчивого штамма в сочетании с мутацией D91N. С учётом отсутствия указанных мутаций у LVX-S-изолятов, комбинированная мутация D91N/Y/G+N87K значимо ассоциирована с устойчивостью клинических изолятов и выявлена у 50,0% (6/12) LVX-R-изолятов (p < 0,001). Следует отметить, что мутации D91N/Y/G, N87K и A88P в гене gyrA были обнаружены исключительно у изолятов, входящих в группу А, и отсутствовали у монорезистентных изолятов. Все мутации в гене gyrB присутствовали у изолятов H. pylori независимо от их фенотипической устойчивости (p > 0,05). Мутации в позициях 86, 97 гена gyrA и 463 гена gyrB, предположительно ассоциированные с резистентностью H. pylori к LVX, в нашей выборке не обнаружены.
Мутации в генах устойчивости к LVX и MTZ у клинических изолятов H. pylori по сравнению с геномом референсного штамма H. pylori 26695, n (%)
Ген (локус) | Аминокислотная замена | LVX-R (n = 12) | LVX-S (n = 31) | MTZ-R (n = 32) | MTZ-S (n = 11) | p |
gyrA (HP_0701) | D91N/Y/G | 4 (33,3) | 0 | 0,0040 | ||
N87K | 2 (16,7) | 0 | 0,0730 | |||
A88P | 1 (8,3) | 0 | 0,2790 | |||
gyrB (HP_0501) | D481E | 3 (25,0) | 8 (25,8) | ~ 1,0000 | ||
R484K | 3 (25,0) | 8 (25,8) | ~ 1,0000 | |||
rdxA (HP_0954) | S108A/P | 7 (21,9) | 0 | 0,1628 | ||
R16C | 7 (21,9) | 0 | 0,1628 | |||
L62V | 6 (18,7) | 0 | 0,3122 | |||
Ter211R_ext* stop lost & splice region | 1 (3,1) | 0 | ~ 1,0000 | |||
Q50*stop | 1 (3,1) | 0 | ~ 1,0000 | |||
W52*stop | 1 (3,1) | 0 | ~ 1,0000 | |||
H97fs | 1 (3,1) | 0 | ~ 1,0000 | |||
S43fs | 1 (3,1) | 0 | ~ 1,0000 | |||
I182fs | 1 (3,1) | 0 | ~ 1,0000 | |||
M120fs | 1 (3,1) | 0 | ~ 1,0000 | |||
R131K | 12 (37,5) | 1 (9,1) | 0,1290 | |||
T31E | 13 (40,6) | 5 (45,4) | 0,7794 | |||
D59N | 30 (93,7) | 9 (81,8) | 0,2665 | |||
frxA (HP_0642) | K18fs | 17 (53,1) | 5 (45,4) | 0,6606 | ||
Y19fs | 1 (3,1) | 0 | ~ 1,0000 | |||
Q27*stop | 1 (3,1) | 0 | ~ 1,0000 | |||
R23fs | 1 (3,1) | 0 | ~ 1,0000 | |||
I44fs | 1 (3,1) | 0 | ~ 1,0000 | |||
A70fs | 1 (3,1) | 0 | ~ 1,0000 | |||
R106fs | 1 (3,1) | 1 (9,1) | 0,4507 | |||
W137*stop | 1 (3,1) | 0 | ~ 1,0000 | |||
Q141*stop | 1 (3,1) | 0 | ~ 1,0000 | |||
V215fs | 10 (31,2) | 6 (54,5) | 0,1679 | |||
fdxB (HP_1508) | N424fs | 1 (3,1) | 1 (9,1) | 0,4507 | ||
K426fs | 1 (3,1) | 1 (9,1) | 0,4507 | |||
fur (HP_1027) | C150Y | 3 (9,4) | 2 (18,2) | 0,5890 | ||
N118Q | 7 (21,9) | 3 (27,3) | 0,6982 |
Всего в гене rdxA выявлено 56 точечных мутаций, из которых 4 (H97fs, S43fs, I182fs, M120fs) относились к мутациям, приводящим к сдвигу рамки считывания (frameshift, fs), 2 (Q50*stop, W52*stop) — к преждевременной терминации рамки считывания, Ter211R_ext* — к потере стоп-кодона и слиянию rdxA с соседним геном HP_0953. Две мутации (S108A/P, L62V) выявлены нами впервые у MTZ-R-изолятов и отсутствовали у MTZ-S-изолятов, однако без ассоциации с фенотипической устойчивостью (p > 0,05). Мутации R131K, T31E и D59N обнаружены у изолятов H. pylori в 2 фенотипических группах и не связаны с фенотипической устойчивостью к MTZ.
В гене frxA нами выявлены 7 мутаций, приводящих к сдвигу рамки считывания, из которых 4 (Y19fs, R23fs, I44fs, A70fs) присутствовали в геноме только MTZ-R-изолятов, 3 (K18fs, R106fs, V215fs) выявлены у изолятов обеих фенотипических групп. В то же время 3 нонсенс-мутации, приводящие к преждевременной терминации белка, обнаружены исключительно в геномах MTZ-R-штаммов (таблица). В гене fdxB 2 мутации (N424fs, K426fs) присутствовали в геноме как MTZ-R-, так и MTZ-S-изолятов. Ни одна из миссенс-мутаций в генах frxA и fdxB не была ассоциирована с фенотипической устойчивостью к MTZ.
Всего в гене fur выявлено 12 миссенс-мутаций, из которых C150Y и N118Q являлись доминирующими, однако ни одна из них не была связана с фенотипической устойчивостью H. pylori к MTZ. Мутаций, приводящих к сдвигу рамки считывания или замене аминокислоты на стоп-кодон, в гене fur у изолятов нашей выборки не обнаружено.
Обсуждение
Неуклонный рост АБР H. pylori во всём мире существенно влияет на эффективность схем эрадикационной терапии. По результатам метаанализа в России за 2011–2020 гг., наиболее значительный рост устойчивости H. pylori отмечен к MTZ (33,95%) и LVX (20,0%) [15]. Однако анализ исследований за 2015–2019 и 2020–2024 гг. выявил снижение LVX-R с 18,3 до 17,1% [15, 16]. Вместе с тем необходимо признать, что данные по уровням и распространённости АБР отсутствуют для большинства регионов России [2].
Наше исследование, включающее 43 клинических изолята H. pylori, выделенных за 2014–2022 гг. на территории Санкт-Петербурга, показало высокий уровень MTZ-R — 74,4%, что указывает на необходимость проведения полномасштабного исследования резистентности H. pylori в данном регионе и ставит под сомнение эффективность применения MTZ в схеме эрадикационной терапии. Напротив, к LVX — препарату 2-й и 3-й линий — штаммы нашей выборки показали более низкую устойчивость — 27,9%. Однако стоит отметить, что среди LVX-R-изолятов лишь 15,0% являлись монорезистентными.
Cуммируя представленные нами ранее данные по устойчивости данной выборки к кларитромицину, 20,9% изолятов H. pylori являются полирезистентными и проявляют устойчивость одновременно к 3 АБП: LVX, MTZ и кларитромицину [17]. Полирезистентные штаммы H. pylori являются глобальной проблемой современной гастроэнтерологии и основным препятствием для эффективного проведения эрадикационной терапии. При наличии резистентности H. pylori к одному из основных АБП частота неудач стандартных схем эрадикации может достигать 30%, а при двойной резистентности — превышать 70% [18, 19]. Поскольку выбор схемы эрадикации носит эмпирический характер, полученные данные диктуют необходимость глобального регионального и локального мониторинга АБР H. pylori в нашей стране, а также адаптации соответствующих стратегий лечения в каждом регионе в зависимости от полученных данных и внедрения программы рационального использования АБП в схемах эрадикационной терапии [20].
В связи с тем, что полногеномное секвенирование является наиболее точным, надёжным, быстрым и эффективным методом определения известных и поиска новых паттернов резистентности, мы использовали данный метод для выявления детерминант резистентности к LVX и MTZ, а также их ассоциации с фенотипической лекарственной устойчивостью российской популяции H. pylori.
Анализ полученных данных показал, что из всех мутаций в генах gyrA и gyrB только D91Y/N/G в гене gyrA значимо ассоциирована с фенотипической устойчивостью H. pylori к LVX (p < 0,05). Другая мутация, связанная с развитием резистентности к фторхинолонам, — N87K — в нашем исследовании выявлена лишь в 16,7% случаев (p > 0,05). Тем не менее, учитывая низкую частоту обнаружения мутации D91Y/N/G, а также отсутствие мутаций D91 и N87 у LVX-S-изолятов, целесообразно рассматривать комбинированную детекцию мутаций D91Y/N/G и N87K в качестве более надёжных предикторов устойчивости H. pylori к LVX. C другой стороны, принимая во внимание высокую долю LVX-R-штаммов, не несущих маркеров резистентности в своём геноме (50,0%), генотипирование гена gyrA для детекции фенотипической устойчивости к LVX может быть недостаточным, что, в свою очередь, ставит под сомнение рациональность использования генотипа D91+N87 gyrA в качестве единственных мишеней при разработке ПЦР-тестов для определения АБР H. pylori в клинической практике. Следует отметить, что мутации D91 и N87 присутствовали геноме только полирезистентных изолятов, тогда как среди монорезистентных к LVZ изолятов данные мутации не выявлены. Это может свидетельствовать как о существовании других, не изученных механизмов резистентности, так и об участии фенотипических механизмов резистентности, таких как изменение уровня экспрессии систем эффлюкса, формирования биоплёнок и др. Кроме того, полирезистентные штаммы могут возникать под давлением комбинированной антибиотикотерапии, что стимулирует накопление мутаций, в том числе в гене gyrA, связанного с устойчивостью к фторхинолонам. Полученные данные нуждаются в дальнейшем изучении с использованием расширенной выборки устойчивых и чувствительных к LVX изолятов H. pylori.
Ввиду высокой гетерогенности штаммов H. pylori изучение и выявление механизмов устойчивости H. pylori к MTZ остаётся сложной задачей. Принято считать, что устойчивость H. pylori к MTZ обусловлена главным образом инактивацией генов rdxA и frxA, кодирующих восстановленную форму никотинамидадениндинуклеотидфосфат (NAD(P)H) нитроредуктазы и флавиннитроредуктазы, катализирующих снижение количества MTZ в клетке [21]. В многочисленных зарубежных исследованиях показано, что большинство MTZ-R-штаммов H. pylori являются носителями множественных нонсенс- и/или fs-мутаций, приводящих к потере функциональных сайтов связывания нитроредуктаз.
В нашем исследовании мутации в гене rdxA, приводящие к потере стоп-кодона, преждевременной терминации или сдвигу рамки считывания, были обнаружены у 21,9% MTZ-R-изолятов и отсутствовали в группе MTZ-S-изолятов. Это может указывать на роль данного гена в развитии АБР у российской популяции H. pylori. Согласно данным E.G. Chua и соавт., мутация R16H/C в гене rdxA ассоциирована с фенотипической устойчивостью изолятов H. pylori к MTZ [22]. Результаты настоящего исследования показали, что мутация R16C, а также мутации S108A/P и L62V в гене rdxA встречались только у изолятов, устойчивых к MTZ, с частотой 21,9, 21,9 и 18,7% соответственно. Однако, учитывая неравномерное распределение долей MTZ-R и MTZ-S изолятов в нашей выборке (74,4 и 25,6%), влияние данных мутаций на развитие фенотипической устойчивости к MTZ ещё предстоит выяснить.
Аналогично, множественные мутации, приводящие к потере функциональности генов frxA и fdxB, могут являться предикторами фенотипической устойчивости H. pylori к MTZ. В нашем исследовании fs-мутации в указанных генах, приводящие к сдвигу рамки считывания, распределялись между изолятами как устойчивой, так и чувствительной фенотипических групп. В гене fur мутаций, ассоциированных с фенотипической устойчивостью к MTZ, не выявлено. Полученные результаты косвенно свидетельствуют о том, что указанные гены, по всей видимости, не играют ключевой роли в развитии устойчивости H. pylori к MTZ в исследуемой популяции. Тем не менее для более точного понимания механизмов резистентности необходимо дополнительное исследование, включающее комплексный анализ других потенциальных генетических факторов в совокупности с возможными синергетическими или эпистатическими взаимодействиями.
Ограничения исследования. Размер выборки ограничен как численно, так и территориально, что не позволяет выявить закономерности генотипической и фенотипической устойчивости изолятов H. pylori на всей территории России. Наблюдалось неравномерное распределение устойчивых и чувствительных к MTZ и LVX изолятов, что существенно осложняет интерпретацию полученных данных и снижает статистическую надёжность результатов. Однако, несмотря на указанные ограничения, наши данные предоставили важную информацию о паттернах резистентности российских клинических изолятов H. pylori, а также подчеркнули необходимость глобального мониторинга АБР в нашей стране.
Заключение
На основании данных полногеномного секвенирования, выполненного нами впервые, было проведено комплексное исследование фенотипической и генотипической резистентности российских клинических изолятов H. pylori к LVX и MTZ. Результаты исследования продемонстрировали высокую долю изолятов, устойчивых к MTZ и LVX, а также высокую частоту полирезистентных изолятов, устойчивых одновременно к 3 АБП (LVX + MTZ + кларитромицин). Несмотря на ограниченный размер выборки, нам удалось подтвердить ключевую роль мутаций D91 и N87 в гене gyrA в формировании устойчивости H. pylori к LVX. Однако высокая частота устойчивых изолятов, не несущих известных детерминант резистентности, ставит под сомнение эффективность существующих и разрабатываемых тест-систем для определения чувствительности H. pylori, а также диктует необходимость проведения более масштабных исследований механизмов АБР изолятов H. pylori.
Полученные результаты подчёркивают исключительную важность вопросов, связанных с непрерывным мониторингом АБР H. pylori в нашей стране, а также контролем динамики изменения генома и механизмов формирования АБР. Это, в свою очередь, может стать предпосылкой к оптимизации схем эрадикационной терапии и повышению эффективности лечения инфекций, обусловленных H. pylori, в разных регионах России.
1 URL: https://github.com/tseemann/snippy
2 URL: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1011037
Об авторах
Дарья Андреевна Старкова
Санкт-Петербургский научно-исследовательский институт эпидемиологии и микробиологии имени Пастера
Email: dariastarkova13@gmail.com
ORCID iD: 0000-0003-3199-8689
канд. биол. наук, с. н. с. лаб. идентификации патогенов, с. н. с. лаб. молекулярной эпидемиологии и эволюционной генетики
Россия, Санкт-ПетербургНикита Сергеевич Гладышев
Российский научный центр хирургии имени академика Б.В. Петровского
Email: krinege@mail.ru
ORCID iD: 0000-0003-2732-5676
н. с. лаб. морфологии и патологии опорно-двигательного аппарата Научно-исследовательского института морфологии человека имени академика А.П. Авцына
Россия, МоскваДмитрий Евгеньевич Полев
Санкт-Петербургский научно-исследовательский институт эпидемиологии и микробиологии имени Пастера
Email: polev@pasteurorg.ru
ORCID iD: 0000-0001-9679-2791
канд. биол. наук, с. н. с., зав. лаб. метагеномных исследований
Россия, Санкт-ПетербургАлина Тимуровна Саитова
Санкт-Петербургский научно-исследовательский институт эпидемиологии и микробиологии имени Пастера
Email: saitova@pasteurorg.ru
ORCID iD: 0000-0002-5921-0745
м. н. с. лаб. метагеномных исследований
Россия, Санкт-ПетербургКсения Александровна Глазунова
Санкт-Петербургский государственный университет
Email: glazunovak03@mail.ru
ORCID iD: 0009-0009-8001-2143
Студентка
Россия, Санкт-ПетербургСветлана Александровна Егорова
Санкт-Петербургский научно-исследовательский институт эпидемиологии и микробиологии имени Пастера
Автор, ответственный за переписку.
Email: egorova@pasteurorg.ru
ORCID iD: 0000-0002-7589-0234
д-р мед. наук, зав. лаб. идентификации патогенов
Россия, Санкт-ПетербургАлена Владимировна Сварваль
Санкт-Петербургский научно-исследовательский институт эпидемиологии и микробиологии имени Пастера
Email: svarval@pasteurorg.ru
ORCID iD: 0000-0001-9340-4132
канд. мед. наук, с. н. с. лаб. идентификации патогенов
Россия, Санкт-ПетербургСписок литературы
- Moss S.F., Shah S.C., Tan M.C., El-Serag H.B. Evolving concepts in Helicobacter pylori management. Gastroenterology. 2024;166(2):267–83. DOI: https://doi.org/10.1053/j.gastro.2023.09.047.
- Ивашкин В.Т., Маев И.В., Лапина Т.Л. и др. Клинические рекомендации Российской гастроэнтерологической ассоциации по диагностике и лечению инфекции Helicobacter pylori у взрослых. Российский журнал гастроэнтерологии, гепатологии, колопроктологии. 2018;28(1):55–70. Ivashkin V.T., Maev I.V., Lapina T.L., et al. Diagnostics and treatment of Helicobacter pylori infection in adults: Clinical guidelines of the Russian gastroenterological association. Russian Journal of Gastroenterology, Hepatology, Coloproctology. 2018;28(1):55–70. DOI: https://doi.org/10.22416/1382-4376-2018-28-1-55-70 EDN: https://elibrary.ru/oulikb
- Malfertheiner P., Megraud F., Rokkas T., et al. European Helicobacter and Microbiota Study group. Management of Helicobacter pylori infection: the Maastricht VI/Florence consensus report. Gut. 2022;71(9):327745. DOI: https://doi.org/10.1136/gutjnl-2022-327745
- Savoldi A., Carrara E., Graham D.Y., et al. Prevalence of antibiotic resistance in Helicobacter pylori: a systematic review and meta-analysis in World Health Organization regions. Gastroenterology. 2018;155(5):1372–82.e17. DOI: https://doi.org/10.1053/j.gastro.2018.07.007
- Zhao M., Zhang Y., Liu S., et al. Eradication of Helicobacter pylori reshapes gut microbiota and facilitates the evolution of antimicrobial resistance through gene transfer and genomic mutations in the gut. BMC Microbiol. 2025;25(1):90. DOI: https://doi.org/10.1186/s12866-025-03823-w
- Hasanuzzaman M., Bang C.S., Gong E.J. Antibiotic resistance of Helicobacter pylori: mechanisms and clinical implications. J. Korean Med. Sci. 2024;39(4):e44. DOI: https://doi.org/10.3346/jkms.2024.39.e44
- Schuetz A.N., Theel E.S., Cole N.C., et al. Testing for Helicobacter pylori in an era of antimicrobial resistance. J. Clin. Microbiol. 2024;62(2):e0073223. DOI: https://doi.org/10.1128/jcm.00732-23
- Dascălu R.I., Bolocan A., Păduaru D.N., et al. Multidrug resistance in Helicobacter pylori infection. Front. Microbiol. 2023;14:1128497. DOI: https://doi.org/10.3389/fmicb.2023.1128497
- Medakina I., Tsapkova L., Polyakova V., et al. Helicobacter pylori antibiotic resistance: molecular basis and diagnostic methods. Int. J. Mol. Sci. 2023;24(11):9433. DOI: https://doi.org/10.3390/ijms24119433
- Saracino I.M., Pavoni M., Zullo A., et al. Next generation sequencing for the prediction of the antibiotic resistance in Helicobacter pylori: a literature review. Antibiotics (Basel). 2021;10(4):437. DOI: https://doi.org/10.3390/antibiotics10040437
- Zhong Z., Zhang Z., Wang J., et al. A retrospective study of the antibiotic-resistant phenotypes and genotypes of Helicobacter pylori strains in China. Am. J. Cancer Res. 2021;11(10):5027–37.
- Gurevich A., Saveliev V., Vyahhi N., Tesler G. QUAST: quality assessment tool for genome assemblies. Bioinformatics. 2013;29(8):1072–5. DOI: https://doi.org/10.1093/bioinformatics/btt086
- Prjibelski A., Antipov D., Meleshko D., et al. Using SPAdes de novo assembler. Curr. Protoc. Bioinformatics. 2020;70(1):e102. DOI: https://doi.org/10.1002/cpbi.102
- Okonechnikov K., Golosova O., Fursov M. Unipro UGENE: a unified bioinformatics toolkit. Bioinformatics. 2012;28(8):1166–7. DOI: https://doi.org/10.1093/bioinformatics/bts091
- Андреев Д.Н., Маев И.В., Кучерявый Ю.А. Резистентность Helicobacter pylori в Российской Федерации: метаанализ исследований за последние 10 лет. Терапевтический архив. 2020;92(11):24–30. Andreev D.N., Maev I.V., Kucheryavyy Yu.A. Helicobacter pylori resistance in the Russian Federation: a meta-analysis of studies over the past 10 years. Therapeutic Archive. 2020;92(11):24–30. DOI: https://doi.org/10.26442/00403660.2020.11.000795 EDN: https://elibrary.ru/gsmibf
- Andreev D.N., Khurmatullina A.R., Maev I.V., et al. Helicobacter pylori antibiotic resistance in Russia: a systematic review and meta-analysis. Antibiotics (Basel). 2025;14(5):524. DOI: https://doi.org/10.3390/antibiotics14050524
- Starkova D., Gladyshev N., Polev D., et al. First insight into the whole genome sequence variations in clarithromycin resistant Helicobacter pylori clinical isolates in Russia. Sci. Rep. 2024;14(1):20108. DOI: https://doi.org/10.1038/s41598-024-70977-4
- Zou Y., Qian X., Liu X., et al. The effect of antibiotic resistance on Helicobacter pylori eradication efficacy: а systematic review and meta-analysis. Helicobacter. 2020;25(4):e12714. DOI: https://doi.org/10.1111/hel.12714
- Gatta L., Scarpignato C., Fiorini G., et al. Impact of primary antibiotic resistance on the effectiveness of sequential therapy for Helicobacter pylori infection: lessons from a 5-year study on a large number of strains. Aliment. Pharmacol. Ther. 2018;47(9):1261–9. DOI: https://doi.org/10.1111/apt.14597
- Полякова В.В., Бодунова Н.А., Цапкова А., Бордин Д.С. Резистентность Helicobacter pylori к антибиотикам и возможности оптимизации эрадикационной терапии. Эффективная фармакотерапия. 2024;20(46):36–44. Polyakova V.V., Bodunova N.A., Tsapkova L.A., Bordin D.S. Helicobacter pylori resistance to antibiotics and possibilities for optimization of eradication therapy. Effective Pharmacotherapy. 2024;20(46):36–44. DOI: https://doi.org/10.33978/2307-3586-2024-20-46-36-44 EDN: https://elibrary.ru/motwfv
- Tuan V.P., Narith D., Tshibangu-Kabamba E., et al. A next-generation sequencing-based approach to identify genetic determinants of antibiotic resistance in Cambodian Helicobacter pylori clinical isolates. J. Clin. Med. 2019;8(6):858. DOI: https://doi.org/10.3390/jcm8060858
- Chua E.G., Debowski A.W., Webberley K.M., et al. Analysis of core protein clusters identifies candidate variable sites conferring metronidazole resistance in Helicobacter pylori. Gastroenterol. Rep. (Oxf.). 2019;7(1):42–9. DOI: https://doi.org/10.1093/gastro/goy048
Дополнительные файлы

